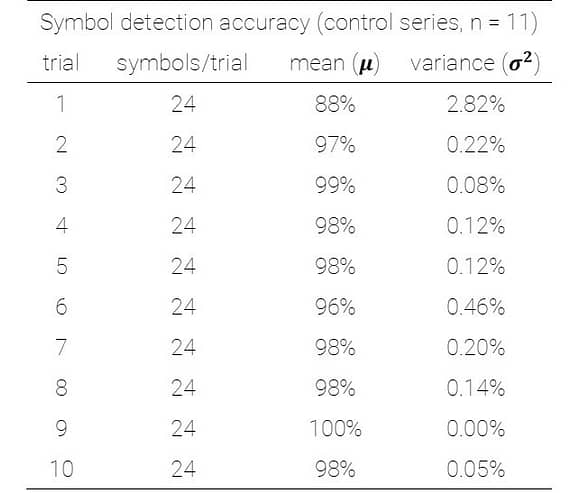
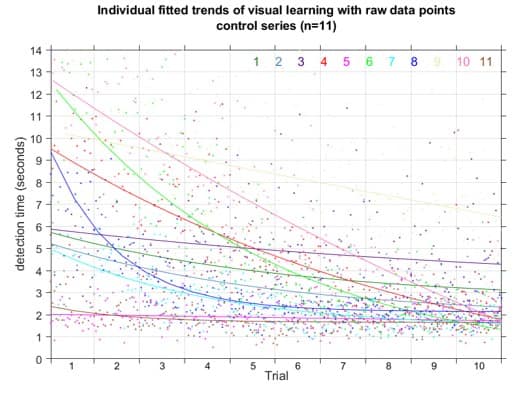
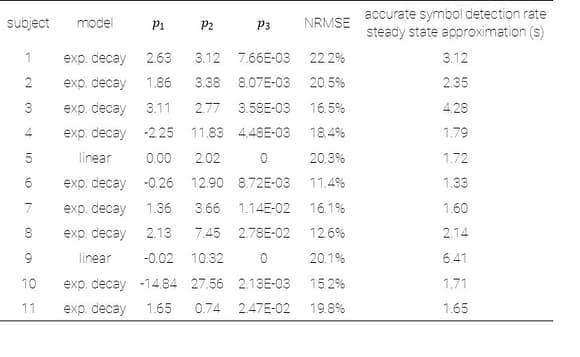
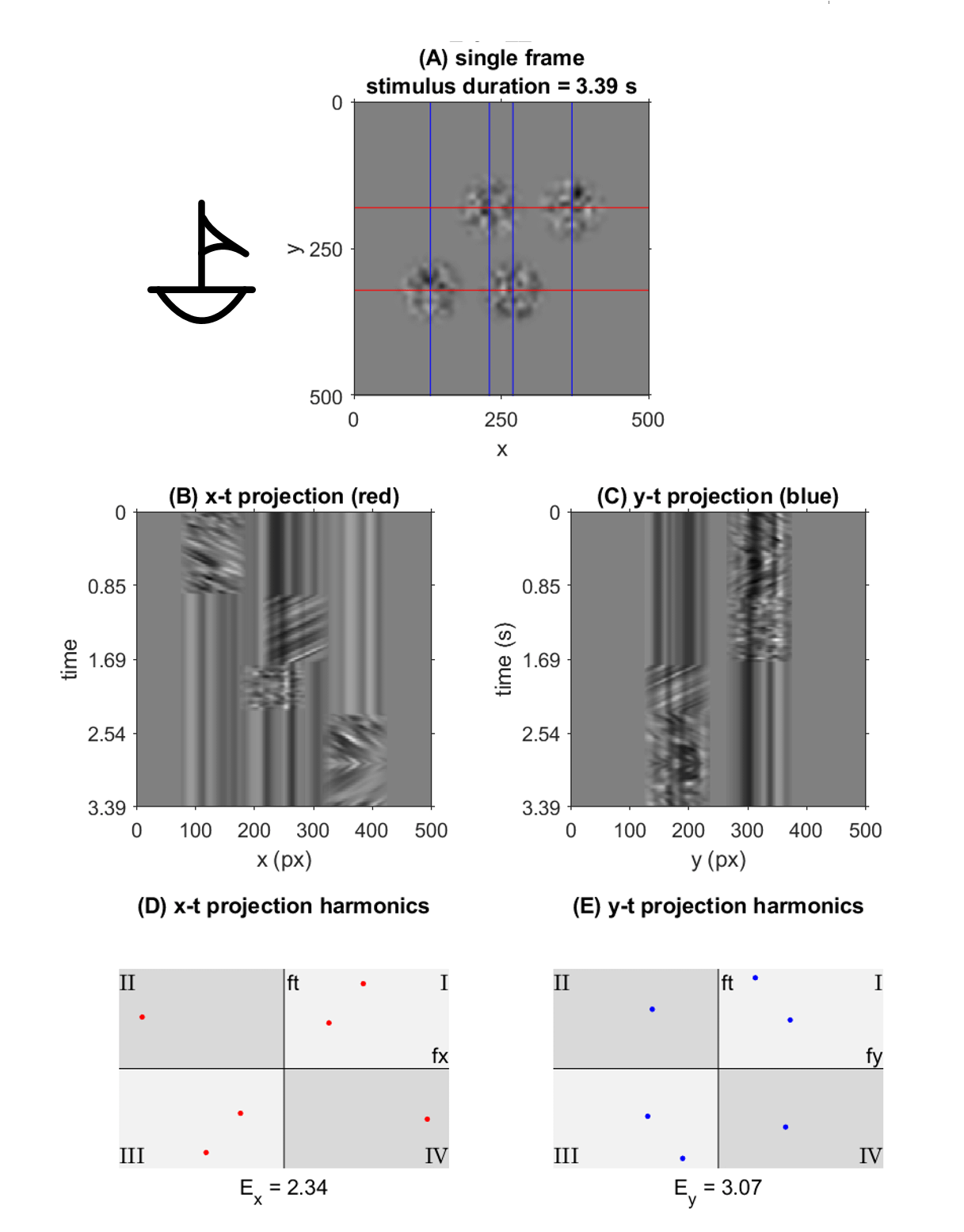
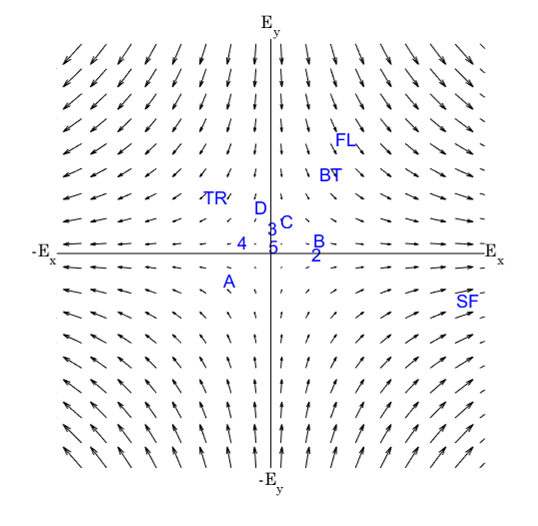
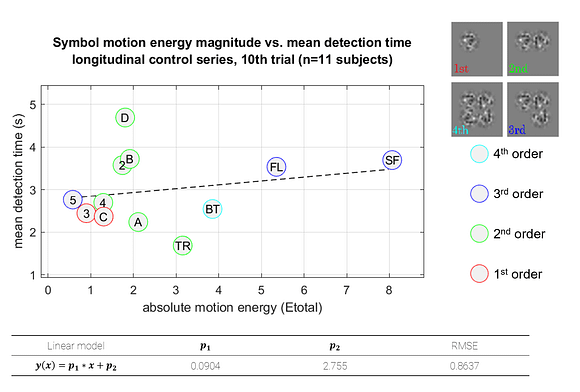
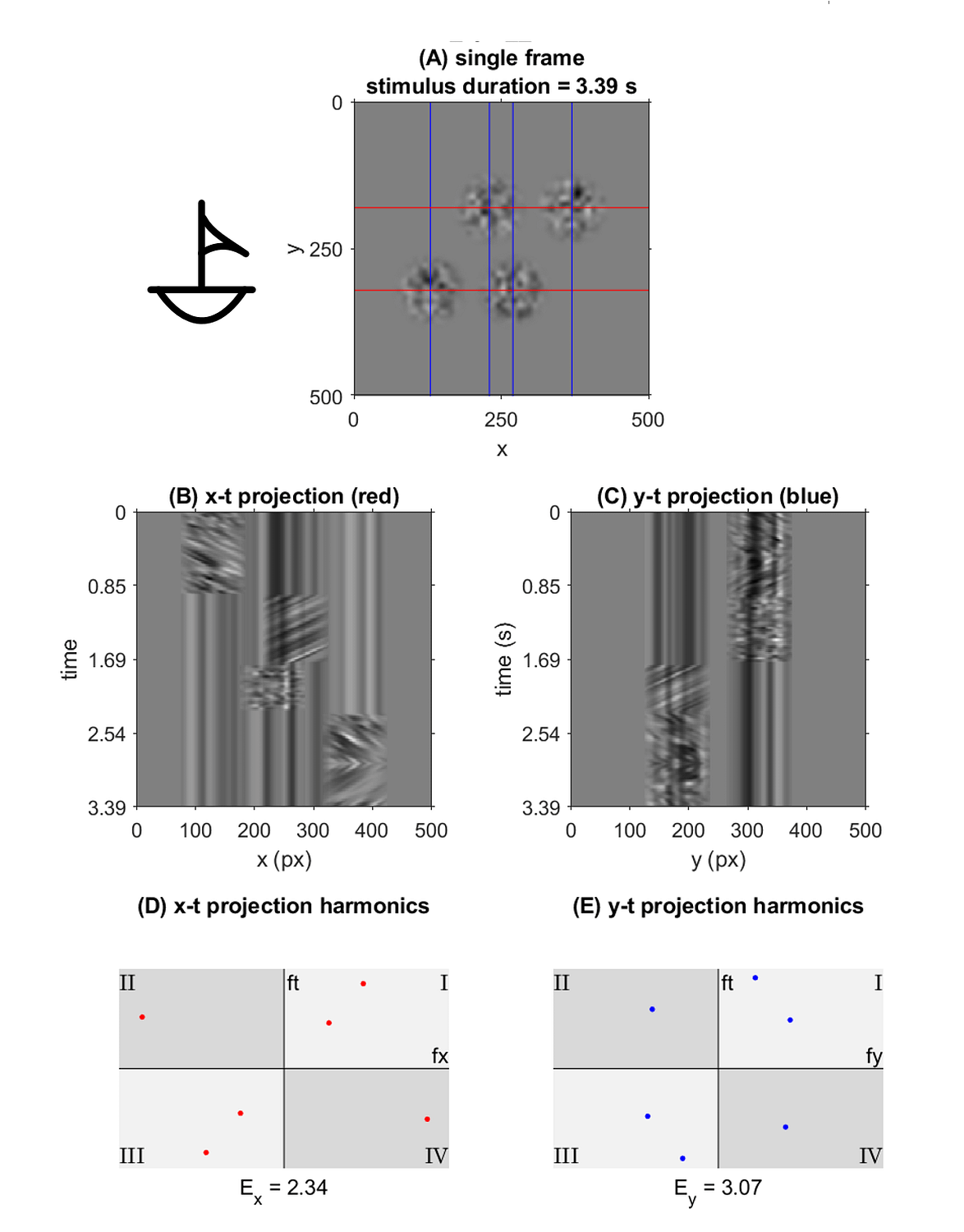
This representation shows the performance of each symbol in detection speed across the control environment versus absolute motion energy. A linear trend is fit to approximate the relationship between absolute motion energy and steady-state detection rate (calculated from the final trial in the control series, n=11). Absolute motion energies of higher complexity symbols such as *stick figure* and *boat* were greater than simpler geometries such as *4* or *C* which is consistent with the expectation that symbols of higher-order (more apertures) and longer duration convey more motion energy. What’s interesting in this representation is a noticeable cluster of recognition times (in seconds) across the majority of symbols presented in the control series. In terms of spatial differentiation of time over motion energy, two significant outliers deviate from the approximate trend. A representation of absolute motion energy versus detection time is shown below, with symbols demarcated by their order (1st through 4th order, indicating the number of apertures present in the codex block).
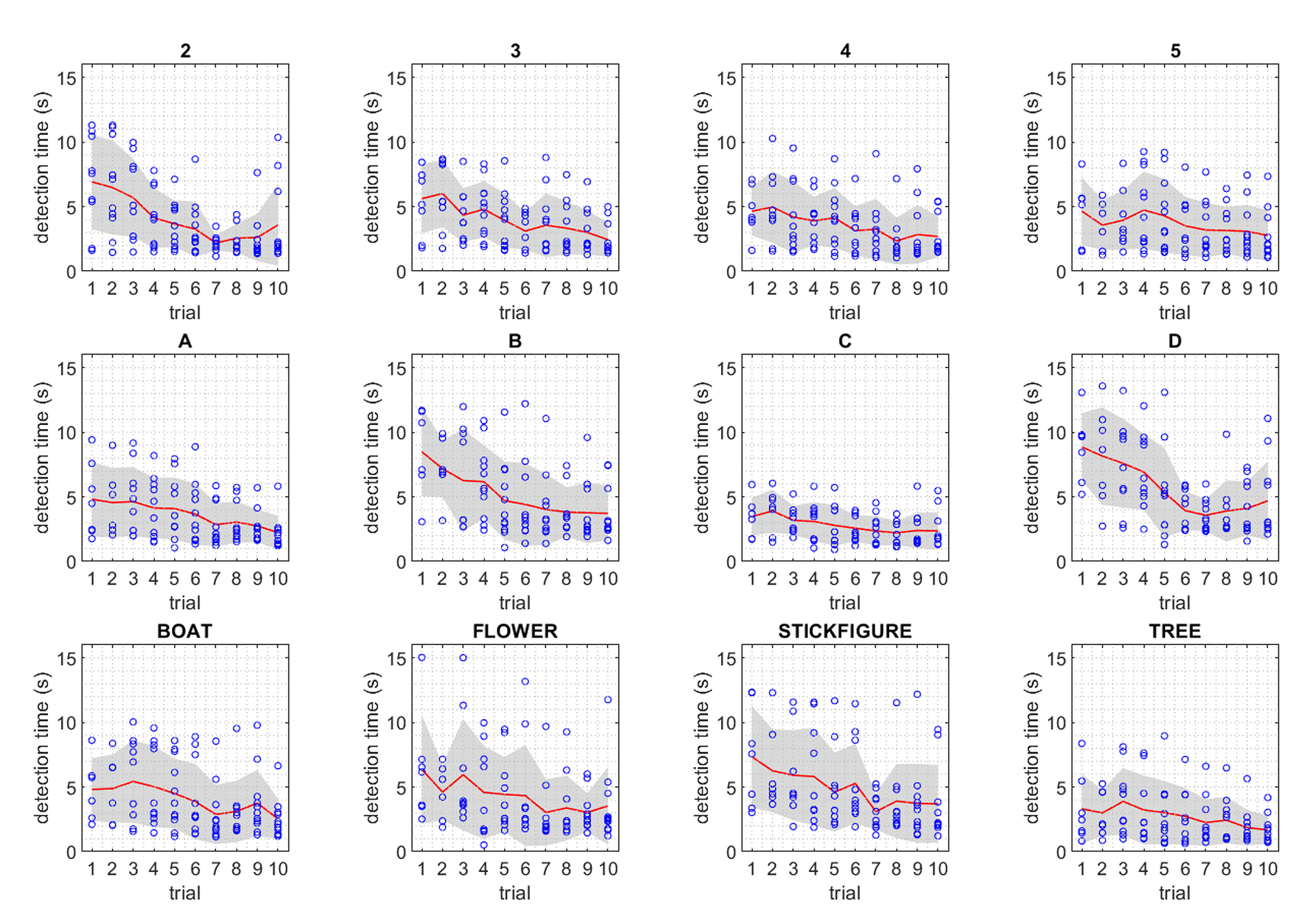
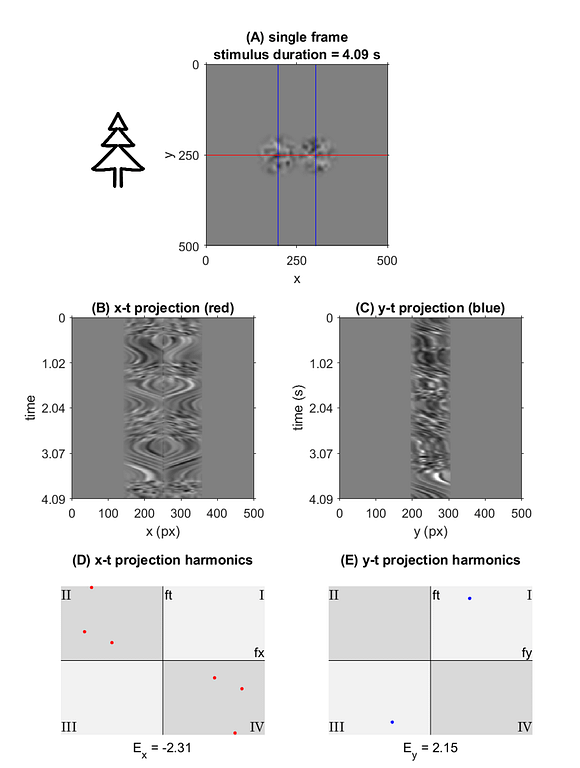
Notable outliers above, *D* and *TR* (shorthand for *tree*), while both 2nd order symbols, fall on either side of the trend. Improvement in detection rates observed for *TR* is best explained by it’s uniqueness in the codex – *TR* was the only symbol to present motion stimulus in multiple apertures simultaneously. Improvements in detection times over repeated trials could be attributed to many factors. One of the most significant is the ability to discriminate symbols in the context of a known subset. In this respect, the uniqueness of *TR* contributed significantly to its low mean detection times.
Uniqueness also played a critical role in the outcomes for symbols *D* and *B*. While having comparatively low motion energies, these required the most time for accurate detection. In terms of stroke trajectories, *D* and *B* are functionally identical for the first half of the symbol presentation. This lack of uniqueness imposes a limit on improvements in detection rates for these symbols. Furthermore, we know from prior work in parameterizing MIPS stimuli, tangential motion, such as the vertical downstroke of *B* and *D* symbols, is far less pronounced than horizontal strokes moving toward or away from the fovea [26], and could have also impacted the perceptibility of these codex blocks.
Uniqueness as a determinant of detection rate does not account for the discrepancy between *B* and *D*, however; to account for this, we must consider the impact of features within white Gaussian noise grain. When grain moves behind an aperture along the stroke path, patterns and features in the grain can create small apparent motion effects. In the case of *D*, an anomaly in the grain causes the downward stroke to appear wavy. These small motion effects were sufficient to cause confusion in the first half-second of symbol perception, greatly limiting improvements in detection time for *D*. For future codex development, one possible approach to identifying unwanted motion effects from grain anomalies could include comparing the desired motion with optical flow measurements.
The role of confusion (i.e. false positives), while not formally evaluated in these experiments, is a critical consideration in the design of a visual language. Within the results of the experimentation described here, there were several recorded instances of false-positive symbol detection between the characters *B* and *D*, but not between any other codex blocks. While there are other instances of similar stroke geometries, such as *3* and *2*, these blocks were distinct in other easily perceptible ways (i.e. number of apertures or codex block duration). Only symbols *B* and *D* were similar across stroke geometry, number of apertures, and codex block duration. In future studies to refine and improve the codex, a key component of optimization will be the design of a library to maximize uniqueness across these three parameters.
Ultimately, the majority of symbols fall within a time frame for complex semantic information delivery that may have a significant impact in industry, both in aerospace and aeronautical instrumentation design, but illustrate a positive trend in the development of generating new stroke geometries to further refine a broader development of codices.